在拟南芥中,已确定有数千个基因在花粉和花粉管中特异表达,但很少有基因与花粉萌发和花粉管生长相关,而且到目前为止发现的参与调控途径的基因几乎都是细胞动力学的正调控因子。然而,由于成熟花粉粒直到暴露在合适的条件下或接触到相容的柱头后才会萌发,因此也必然存在抑制花粉萌发的负面调控机制。1. 为了揭示正常花粉萌发所需的调控机制,研究者筛选了在花粉和花粉管中特异表达的基因。研究者发现了一个未知的拟南芥基因,该基因编码一个51.4kD的蛋白,含有7个WD40重复序列,被命名为 “金箍棒” (JINGUBANG, JGB),它在确保花粉正常萌发方面发挥着关键作用。研究者为了确定其表达模式,构建了JGB与GUS的融合蛋白并进行了下图所示实验。a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φB:研究人员使用了RT-PCR(左)和gus基因检测法(右)检测其空间特异性表达情况。a,花序;b,花;c,花药;d,花粉管。C:gus基因检测法检测在花粉发育过程中JGB的表达。UNM,单核小孢子;BCP,双细胞花粉;TCP,三细胞花粉。下列说法正确的是( )。A: GAPDH起到了内参基因的作用;JGB只在花器官中特异性表达B: 还需要突变体表型分析证实JGB不是植物生长或繁殖需要的花粉特异基因,该基因才符合筛选标准C: 在C图中,GUS同Hoechst均可以显示目标基因的积累情况D: 在野生型花粉的发育过程中JGB蛋白积累,该蛋白在花粉管生长中具有较强的GUS活性2. 为了探究JGB在花粉萌发过程中所起到的作用,研究者构建了JGB的突变体jgb品系,并进行了下列工作。a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φ图A:两种花粉在添加有不同物质的人工培养基中的萌发率。图B:在不同时间内花粉的萌发率。下列说法正确的是( )。A: 由图片可知,JGB在花粉萌发过程中起到促进的作用B: 钙、镁、钾、硼等矿质元素为正常花粉萌发所必需的C: JGB的作用在于抑制异常活跃的花粉萌发D: 这些结果表明,敲除JGB降低了花粉萌发需求,诱导花粉快速萌发3. 研究者在植株中超表达JGB,得到高水平表达JGB的JGB OX品系。a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φ图A:JGB OX品系的构建过程。图B:以微管蛋白为内参,通过免疫印迹法检测Col-0中JGB的水平,JGB中JGB的水平以及JGB OX中JGB和JGB-GFP的水平。图C、D:三种不同基因型花粉在人工培养基上萌发情况和萌发率。图E:表现出较弱JGB-GFP荧光的JGB OX花粉通常会萌发。图F、G、H:人工授粉后,不同花粉在柱头上的萌发情况和萌发率。OX-LOW:低表达;OX-HIGH:高表达下列说法错误的是( )。A: 绿色荧光表达水平高的花粉粒,其萌发受抑制的程度也越大B: B图显示与GFP融合的JGB已经成功超表达C: 可使用学生氏t检验比较花粉萌发率的差异D: 在任何情况下,JGB基因对花粉萌发都具有很强的抑制作用E: JGB对花粉萌发的抑制作用可以通过柱头授粉来减弱4. 为了探究JGB基因在花粉萌发中可能的其他作用,研究者进一步进行了以下工作。a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φa d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φ表一:研究人员使用两个杂交实验研究突变对雌雄配子育性的影响。TE:配子传代效率,rm TE=jgb/JBG杂合种子数目:Col-0种子数目。图A、B:授粉后JGB花粉管在柱头上的生长情况,箭头示花粉管末端延伸的距离。图C:杂交产生的角果中各部位产生的JGB子代所占的百分比。jgb突变对配子传代效率的直接影响是( )。A: 提高雄性配子传代效率B: 降低雄性配子传代效率C: 提高雌性配子传代效率D: 降低雌性配子传代效率5. 根据以上图表信息和上题结论,下列说法错误的是( )。A: 在表一中,可使用PCR的方式检测产生的种子的基因型B: 在rm 8h时,jgb和野生型的花粉管长度差异不显著,表示JGB基因突变对花粉管的延伸影响不大C: JGB基因突变对花粉的萌发初期没有影响,但是4至rm 6h的花粉管生长速度降低D: jgb花粉管在到达子房下半部时生长受影响6. 研究者利用JGB-GFP融合蛋白研究JGB的定位。a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φA:三细胞花粉的共聚焦显微镜图像。显示JGB-GFP荧光在花粉中的定位。B:同一个四分子不同发育时期的图像。使用JGB/JGB-GFP品系。UNM,单核小孢子;BCP,双细胞花粉;TCP,三细胞花粉。C:花粉管萌发过程中的图像。pt,花粉管;sn,精子核;vn,营养核。则下列说法错误的是( )。A: JGB主要富集于三细胞花粉中的一个核中B: JGB主要积累于花粉管的精子核内C: 图B显示,JGB-GFP荧光在三细胞花粉期开始出现D: JGB的亚细胞定位在花粉萌发和花粉管生长过程中很可能没有改变E: 可以推测,JGB含有NLS序列7. 研究人员发现,JGB具有NES(核输出)序列,故在细胞核和细胞质中皆有定位。为了研究其具体功能定位于细胞核还是细胞质,进行了下列实验。a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φA至C:研究人员构建了NES序列的突变体品系jgb rm RES^AEV。B,绿色荧光示JGB-GFP融合蛋白的分布。C为B的量化结果。Nu,细胞核;Cy;细胞质。D:在体内实验中,不同花粉的萌发率。E:花粉管生长情况。F为E的量化结果。由以上结果,可以得出结论:控制花粉萌发和花粉管生长的很可能是( )。A: 细胞核JGBB: 细胞质JGB8. 研究人员发现,JGB的作用可能与花粉中茉莉酸的代谢有关。为进一步分析JGB的作用,研究者做了以下工作。a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φA:研究人员使用100-10000 μM的MeJA处理野生型花粉,研究其对野生型花粉萌发率的影响。B为A的量化结果。C、D:探究MeJA和二乙基二硫代氨基甲酸(DIECA,一种JA生物合成的抑制剂)对花粉萌发的影响。E:在不同的品系中与茉莉酸合成有关基因的表达水平。F:不同品系的花粉中茉莉酸类物质的含量。下列说法正确的是( )。A: JA对花粉的萌发起双向调节作用,过高浓度会抑制其萌发B: 在正常生理浓度下,JA对花粉萌发的作用以促进为主C: E图采用了RNA-seq的方法对基因表达水平进行检测;F图可以使用超高效液相色谱串联三重四级杆质谱(UPLC-triple quadrupole MS)的方式检测JA含量D: JGB基因可以抑制内源JA合成,从而控制花粉萌发9. 为了确定JGB如何影响JA的合成,研究人员筛选了拟南芥转录因子的cDNA文库,找到了四个TCP家族成员——TCP4、TCP8、TCP20和TCP22,作为可能与JGB相互作用的候选蛋白。四种蛋白对JA合成有关基因均起激活作用。为了确认JGB与候选蛋白之间的相互作用,研究者将待选蛋白与转录激活结构域融合并插入pDEST22质粒(含Trp合成基因);并将JGB与GAL4的DNA结合结构域融合并插入pDEST32质粒(含Leu合成基因)。两质粒通过电穿孔法转入酵母AH109品系(有Leu、Trp、His的营养缺陷,并存有UAS序列激活的His基因),将酵母培养至SD/-2培养基(Leu、Trp缺失的选择培养基)和SD/-3(Leu、Trp、His缺失的选择培养基),并观察培养结果,如下图:a d-|||-Stage.12-|||-" tan theta lt 13-|||-Stage44-|||-b-|||-C-|||-UNM Early BCP Late BCP TCP-|||-φA:研究人员使用酵母双杂交检验JGB与TCP-4有无相互作用,结果如图所示。注意:SD/-3的酵母培养基经过了稀释涂布平板处理,自左至右浓度逐步下降B:JGB与TCP-4的GST-pull down结果。SD/-2:SD/-Leu-Trp;SD/-3:SD/-Leu-Trp-His;SD为酵母缺陷型。下列说法正确的是( )。A: 酵母双杂交有较高的假阳性率,所以必须做阴性对照实验。B: A图中第二三横行为阴性对照,SD/-2为阳性对照。C: 图B的主要目的在于验证图A结论,避免假阳性。D: AB图证明,JGB可以与TCP4结合并抑制其活性。E: 图B中使用的技术一般用来检测蛋白质之间在体内的相互作用
在拟南芥中,已确定有数千个基因在花粉和花粉管中特异表达,但很少有基因与花粉萌发和花粉管生长相关,而且到目前为止发现的参与调控途径的基因几乎都是细胞动力学的正调控因子。然而,由于成熟花粉粒直到暴露在合适的条件下或接触到相容的柱头后才会萌发,因此也必然存在抑制花粉萌发的负面调控机制。
1. 为了揭示正常花粉萌发所需的调控机制,研究者筛选了在花粉和花粉管中特异表达的基因。研究者发现了一个未知的拟南芥基因,该基因编码一个51.4kD的蛋白,含有7个WD40重复序列,被命名为 “金箍棒” (JINGUBANG, JGB),它在确保花粉正常萌发方面发挥着关键作用。研究者为了确定其表达模式,构建了JGB与GUS的融合蛋白并进行了下图所示实验。
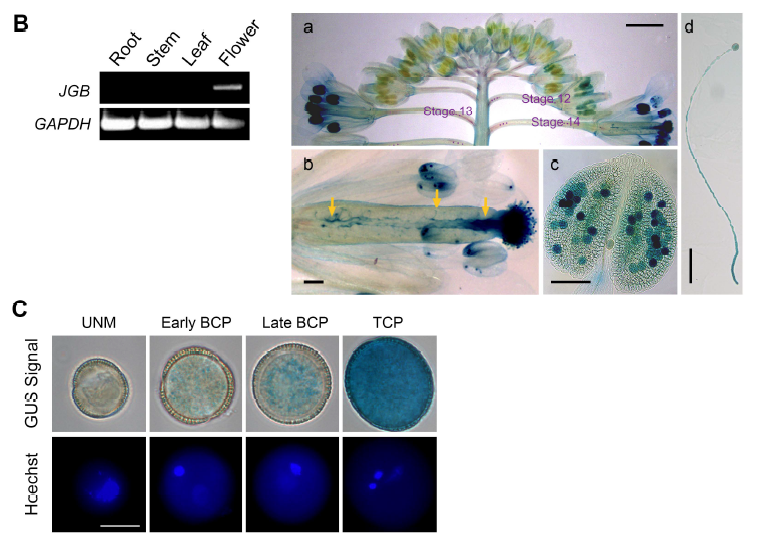
B:研究人员使用了RT-PCR(左)和gus基因检测法(右)检测其空间特异性表达情况。
a,花序;b,花;c,花药;d,花粉管。
C:gus基因检测法检测在花粉发育过程中JGB的表达。UNM,单核小孢子;BCP,双细胞花粉;TCP,三细胞花粉。
下列说法正确的是( )。
A: GAPDH起到了内参基因的作用;JGB只在花器官中特异性表达
B: 还需要突变体表型分析证实JGB不是植物生长或繁殖需要的花粉特异基因,该基因才符合筛选标准
C: 在C图中,GUS同Hoechst均可以显示目标基因的积累情况
D: 在野生型花粉的发育过程中JGB蛋白积累,该蛋白在花粉管生长中具有较强的GUS活性
2. 为了探究JGB在花粉萌发过程中所起到的作用,研究者构建了JGB的突变体jgb品系,并进行了下列工作。
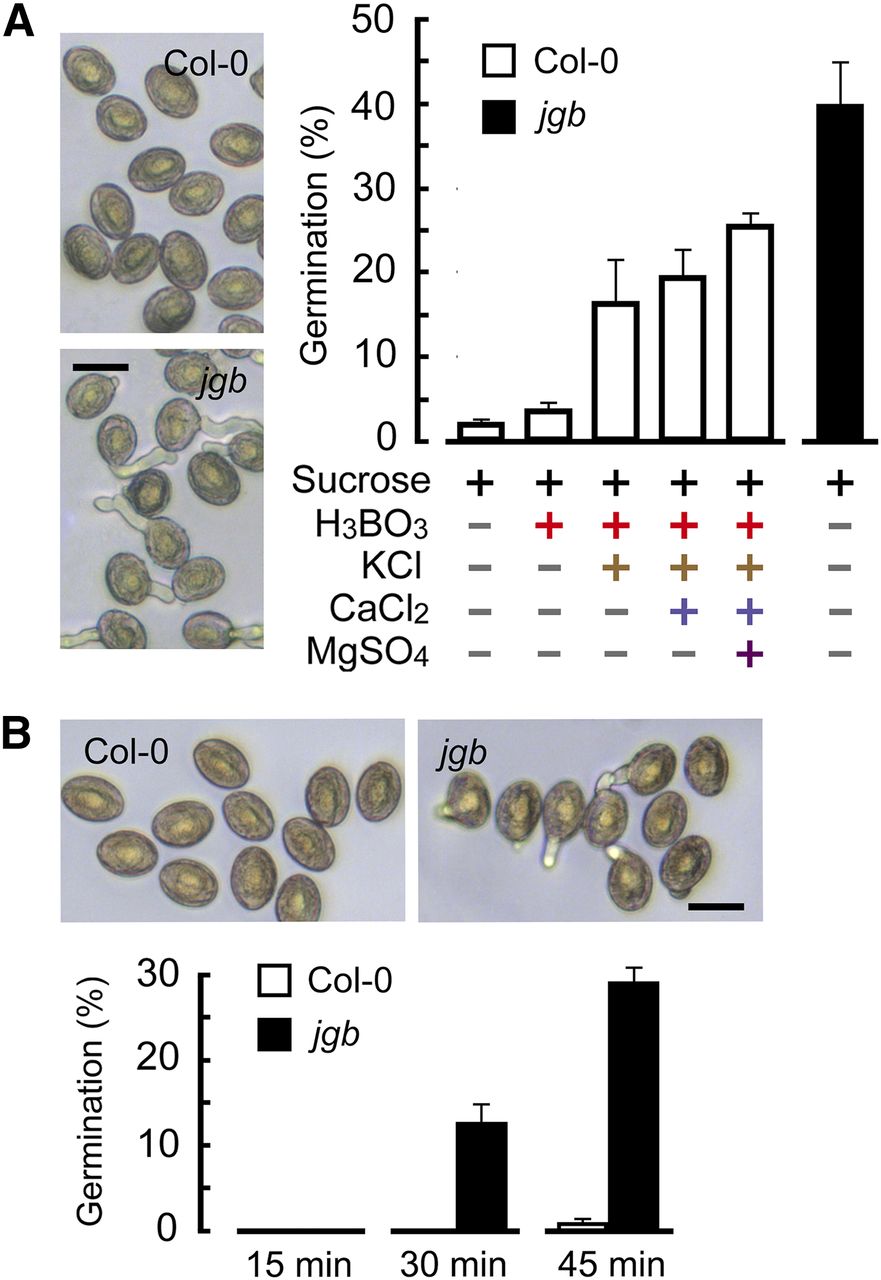
图A:两种花粉在添加有不同物质的人工培养基中的萌发率。
图B:在不同时间内花粉的萌发率。
下列说法正确的是( )。
A: 由图片可知,JGB在花粉萌发过程中起到促进的作用
B: 钙、镁、钾、硼等矿质元素为正常花粉萌发所必需的
C: JGB的作用在于抑制异常活跃的花粉萌发
D: 这些结果表明,敲除JGB降低了花粉萌发需求,诱导花粉快速萌发
3. 研究者在植株中超表达JGB,得到高水平表达JGB的JGB OX品系。
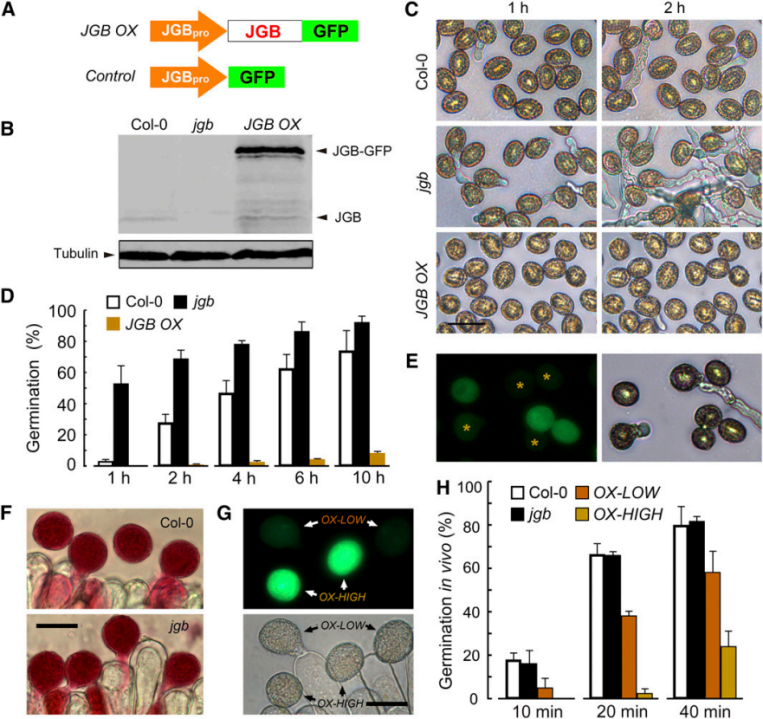
图A:JGB OX品系的构建过程。
图B:以微管蛋白为内参,通过免疫印迹法检测Col-0中JGB的水平,JGB中JGB的水平以及JGB OX中JGB和JGB-GFP的水平。
图C、D:三种不同基因型花粉在人工培养基上萌发情况和萌发率。
图E:表现出较弱JGB-GFP荧光的JGB OX花粉通常会萌发。
图F、G、H:人工授粉后,不同花粉在柱头上的萌发情况和萌发率。OX-LOW:低表达;OX-HIGH:高表达
下列说法错误的是( )。
A: 绿色荧光表达水平高的花粉粒,其萌发受抑制的程度也越大
B: B图显示与GFP融合的JGB已经成功超表达
C: 可使用学生氏t检验比较花粉萌发率的差异
D: 在任何情况下,JGB基因对花粉萌发都具有很强的抑制作用
E: JGB对花粉萌发的抑制作用可以通过柱头授粉来减弱
4. 为了探究JGB基因在花粉萌发中可能的其他作用,研究者进一步进行了以下工作。
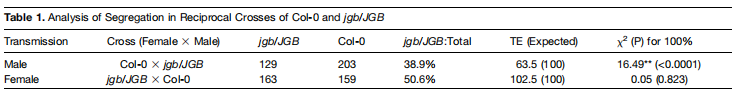
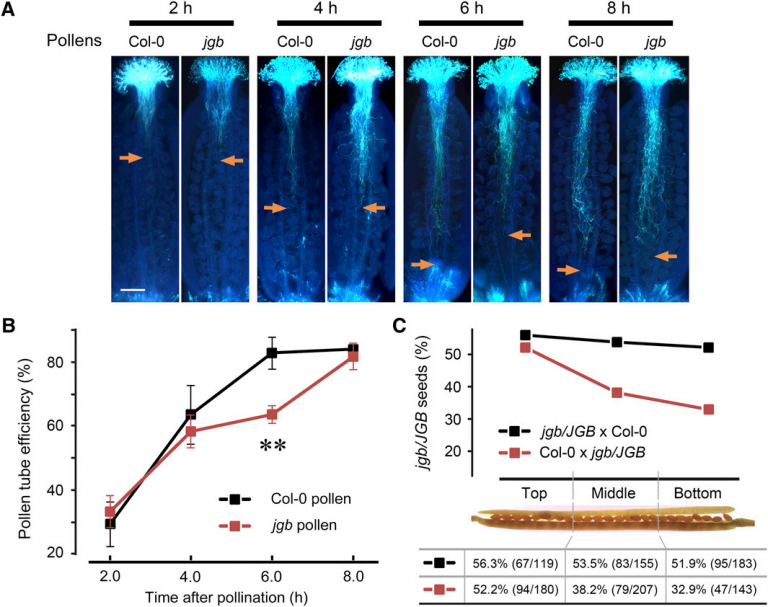
表一:研究人员使用两个杂交实验研究突变对雌雄配子育性的影响。TE:配子传代效率,$$\rm TE=jgb/JBG杂合种子$$数目:Col-0种子数目。
图A、B:授粉后JGB花粉管在柱头上的生长情况,箭头示花粉管末端延伸的距离。
图C:杂交产生的角果中各部位产生的JGB子代所占的百分比。
jgb突变对配子传代效率的直接影响是( )。
A: 提高雄性配子传代效率
B: 降低雄性配子传代效率
C: 提高雌性配子传代效率
D: 降低雌性配子传代效率
5. 根据以上图表信息和上题结论,下列说法错误的是( )。
A: 在表一中,可使用PCR的方式检测产生的种子的基因型
B: 在$$\rm 8h$$时,jgb和野生型的花粉管长度差异不显著,表示JGB基因突变对花粉管的延伸影响不大
C: JGB基因突变对花粉的萌发初期没有影响,但是4至$$\rm 6h$$的花粉管生长速度降低
D: jgb花粉管在到达子房下半部时生长受影响
6. 研究者利用JGB-GFP融合蛋白研究JGB的定位。
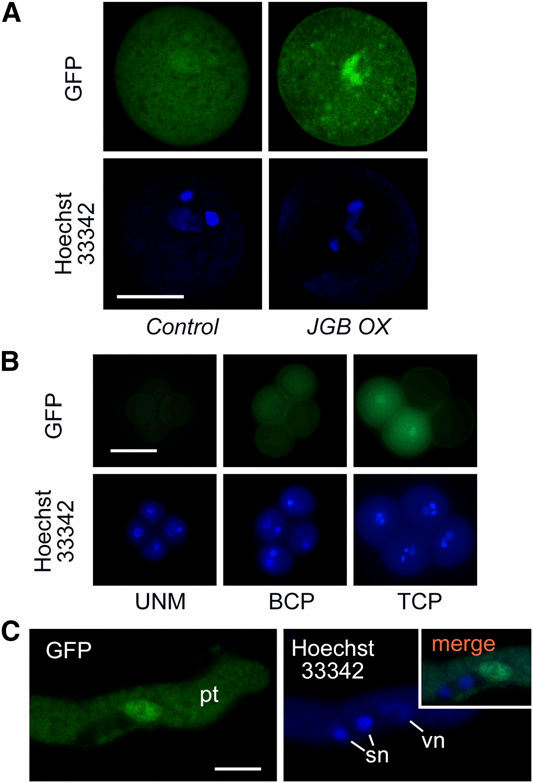
A:三细胞花粉的共聚焦显微镜图像。显示JGB-GFP荧光在花粉中的定位。
B:同一个四分子不同发育时期的图像。使用JGB/JGB-GFP品系。UNM,单核小孢子;BCP,双细胞花粉;TCP,三细胞花粉。
C:花粉管萌发过程中的图像。pt,花粉管;sn,精子核;vn,营养核。
则下列说法错误的是( )。
A: JGB主要富集于三细胞花粉中的一个核中
B: JGB主要积累于花粉管的精子核内
C: 图B显示,JGB-GFP荧光在三细胞花粉期开始出现
D: JGB的亚细胞定位在花粉萌发和花粉管生长过程中很可能没有改变
E: 可以推测,JGB含有NLS序列
7. 研究人员发现,JGB具有NES(核输出)序列,故在细胞核和细胞质中皆有定位。为了研究其具体功能定位于细胞核还是细胞质,进行了下列实验。
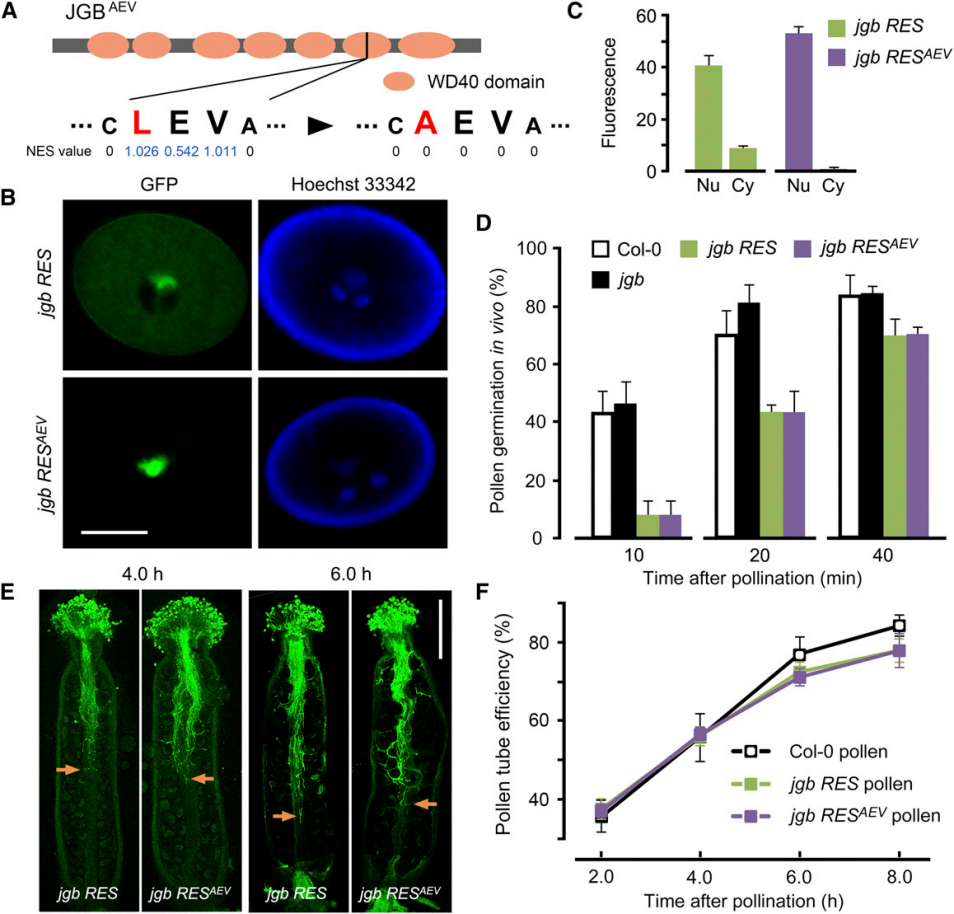
A至C:研究人员构建了NES序列的突变体品系jgb $$\rm RES^{AEV}$$。B,绿色荧光示JGB-GFP融合蛋白的分布。C为B的量化结果。Nu,细胞核;Cy;细胞质。
D:在体内实验中,不同花粉的萌发率。
E:花粉管生长情况。F为E的量化结果。
由以上结果,可以得出结论:控制花粉萌发和花粉管生长的很可能是( )。
A: 细胞核JGB
B: 细胞质JGB
8. 研究人员发现,JGB的作用可能与花粉中茉莉酸的代谢有关。为进一步分析JGB的作用,研究者做了以下工作。
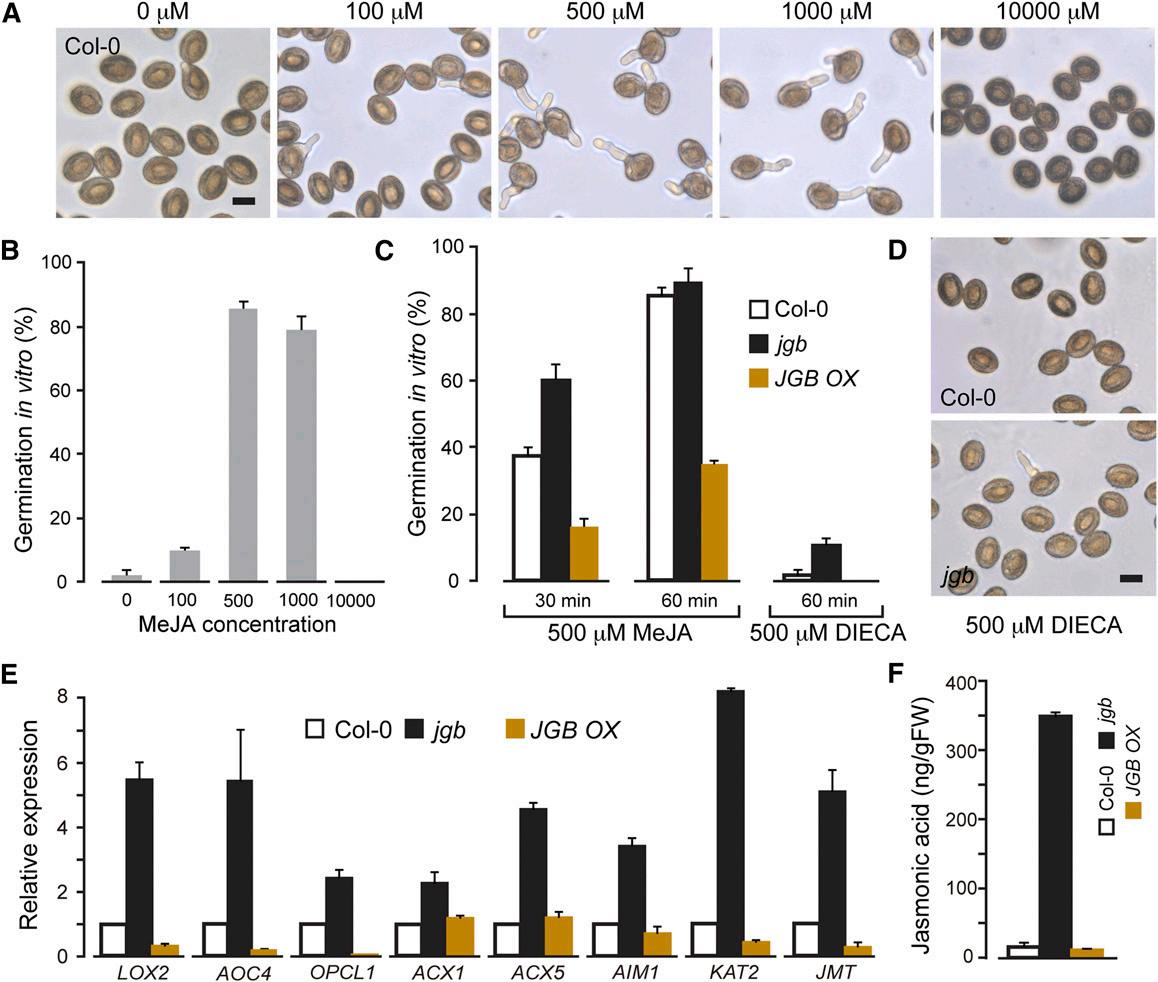
A:研究人员使用100-10000 μM的MeJA处理野生型花粉,研究其对野生型花粉萌发率的影响。B为A的量化结果。
C、D:探究MeJA和二乙基二硫代氨基甲酸(DIECA,一种JA生物合成的抑制剂)对花粉萌发的影响。
E:在不同的品系中与茉莉酸合成有关基因的表达水平。
F:不同品系的花粉中茉莉酸类物质的含量。
下列说法正确的是( )。
A: JA对花粉的萌发起双向调节作用,过高浓度会抑制其萌发
B: 在正常生理浓度下,JA对花粉萌发的作用以促进为主
C: E图采用了RNA-seq的方法对基因表达水平进行检测;F图可以使用超高效液相色谱串联三重四级杆质谱(UPLC-triple quadrupole MS)的方式检测JA含量
D: JGB基因可以抑制内源JA合成,从而控制花粉萌发
9. 为了确定JGB如何影响JA的合成,研究人员筛选了拟南芥转录因子的cDNA文库,找到了四个TCP家族成员——TCP4、TCP8、TCP20和TCP22,作为可能与JGB相互作用的候选蛋白。四种蛋白对JA合成有关基因均起激活作用。
为了确认JGB与候选蛋白之间的相互作用,研究者将待选蛋白与转录激活结构域融合并插入pDEST22质粒(含Trp合成基因);并将JGB与GAL4的DNA结合结构域融合并插入pDEST32质粒(含Leu合成基因)。两质粒通过电穿孔法转入酵母AH109品系(有Leu、Trp、His的营养缺陷,并存有UAS序列激活的His基因),将酵母培养至SD/-2培养基(Leu、Trp缺失的选择培养基)和SD/-3(Leu、Trp、His缺失的选择培养基),并观察培养结果,如下图:
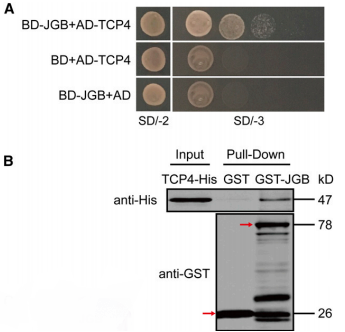
A:研究人员使用酵母双杂交检验JGB与TCP-4有无相互作用,结果如图所示。
注意:SD/-3的酵母培养基经过了稀释涂布平板处理,自左至右浓度逐步下降
B:JGB与TCP-4的GST-pull down结果。
SD/-2:SD/-Leu-Trp;SD/-3:SD/-Leu-Trp-His;SD为酵母缺陷型。
下列说法正确的是( )。
A: 酵母双杂交有较高的假阳性率,所以必须做阴性对照实验。
B: A图中第二三横行为阴性对照,SD/-2为阳性对照。
C: 图B的主要目的在于验证图A结论,避免假阳性。
D: AB图证明,JGB可以与TCP4结合并抑制其活性。
E: 图B中使用的技术一般用来检测蛋白质之间在体内的相互作用
题目解答
答案
1. AB
正确率: 16%, 易错项: C
2. BCD
正确率: 10%, 易错项: A
3. D
正确率: 29%, 易错项: C
4. B
正确率: 46%, 易错项: A
5. B
正确率: 45%, 易错项: C
6. BC
正确率: 15%, 易错项: D
7. A
正确率: 75%, 易错项: B
8. ABD
正确率: 9%, 易错项: C
9. ABCD